Human development is the process where a single cell, a rather unremarkable cell except for perhaps its size, slowly changes into trillions of different-looking cells. Embryology is the study of the early phases of development, and its where all the interesting stuff happens. A round cell, just a ball with no sides and no special parts, grows into an organism with different organs in different places. The image to the left is our Sun. You see it every day, can you tell whether I have the photo right-side up? Is there even a right-side up? Dangit, I'm an embryologist, not an astronomer! All I know is the Sun looks a bit like a fertilized egg, I can't see a top or bottom, I don't see any continents or oceans. An egg has a similar amorphous appearance, but it will morph into the embryo shown in the image below, which has a head-end and tail-end, and clear beginnings of arms and legs.
Human embryo 7 weeks, by GoldenBear is liscensed CC BY SA 3.0
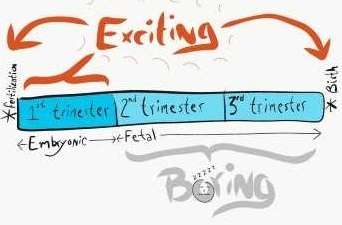
Pregnancy is divided into three equal time periods named trimesters. During the first trimester, a single cell with no shape develops into an embryo, which has shapes. The embryo keeps growing new shapes until it has all of the basic shapes a human needs. At that time we call it a fetus, and the first trimester has ended. The next two trimesters are relatively boring, the fetus grows bigger and the shapes mature, but nothing terribly exciting happens until birth. That's why this is a Histology and Embryology book, not Fetology.
Filmmaker Niko von Glasow, is liscensed CC BY SA 2.0
As human cells divide during the first trimester, they make decisions as to their cell fate, which ultmately determines the number and location of our arms and legs, our head and heart. Errors in these decisions lead to congenital disorders, or birth defects (but see below). These can happen without known cause. Or, certain chemicals can interfere with developmental decisions, and we call these chemicals teratogens. One of the more famous teratogens of my generation is the drug thalidomide, which-- if taken by pregnant women at the wrong time-- interferes with the decision-making processes an embryo uses to decide where to grow limbs. This causes phocomelia, or malformation of the limbs. There are trillions of decisions made between the single-cell stage and birth, errors will occur. Some errors are minor, changing the appearance but not the function of an organ, and we might refer to that as a birth mark, or just human variation. Other mistakes are more severe, and we refer to those as congenital disorders, preferably not birth defects. Patients should not feel their medical professionals think of them as defective, otherwise they seek less health care.
Patterns
This section covers the basic processes that occur during development. Often, more than one will be occuring in the same place at the same time. However, it is useful to try to think about the following changes separately becuase they are controlled by different signals.
Cloning
Mitosis produces clones, or identical cells with identical DNA. However, cells need to differentiate and begin expressing different genes to become different tissues and organs. In the first stages of development cells simply divide and produce more identical cells. Very soon, cells begin to make decisions as to what they are going to become, and these decisions are co-ordinated with other cells, otherwise an embryo could wind up with two butts and no head.
Induction
One cell may signal to another cell and induce (instructs) that cell to differentiate. Chemicals that can induce another cell to change its cell fate are called morphogens (or sometimes called Growth Factors). Morphogens are similar to neurotransmitters or hormones, they often bind to cell-surface receptor proteins which leads to activation of specific transcription factors to alter the expression of different genes, causing the cell to look and function differently. Morphogens can act over different ranges:
Morphogens are usually secreted into ground substance. The amount of morphogen molecules produced, the relative stickiness of the ground substance, the distance away from the source and the number of receptor proteins on the target cell determine how much of a signal a cell receives. If the level of the morphogen signal reaches a certain theshold, it dictates changes to cell fate. Morphogens frequently bind to cell-surface receptor proteins. Activation of these receptors triggers a cascade of signals inside the cell, which ultimately leads to activation of transcription factors, which trigger the transcription of new genes or shut off transcription of genes that are no longer needed. During early development, the genes that are activated or inhibited control cell fate, such as homeobox genes. Homeobox genes are themselves transcription factors that specify which part of the body a particular cell is now a part of, such as part of an arm or part of the upper jaw. They do not encode the proteins that cells use to become part of a specific organ. Instead, homeobox genes allow other genes to be activated later, setting the stage for future changes. Therefore, during early development cells are changing fate, but these changes are not visible. Later in development, morphogens activate genes that encode the proteins that drive differentiation of cells, which leads to visible changes in morphology.
Some morphogens block other cells from producing the same morphogen. When two such morphogens are produced in the same area, complex patterns arise. In the image to the left, blue morphogen induces one cell to differentiate into a blue cell, but blocks neighboring cells from becoming blue cells. These neighbors would be free to produce red morphogen, however, and differentiate into red cells. At the same time, a distant cell produces red morphogen, which induces it to differentiate into a red cell, but blocks its neighbors from doing so. Its neighbors would be free to produce blue morphogen and differentiate into blue cells. The initial pattern that arises will be rings of alternating rings of red and blue cells, or blue and red cells. However, there are regions of red-next-to-red and blue-next-to-blue, which starts a battle. To minimize the number of red-next-to-red and blue-next-to-blue, cells change their cell fate until a stable pattern arises, in this case stripes. If these were pigment-producing cells in a zebra, we'd have a zebra with stripes (the best type of zebra). But this pattern could also be alternating regions of oral mucosa that grow a tooth bud or don't grow a tooth bud, ensuring each tooth bud will have enough space to grow (if yor really want to go down a rabbit-hole, these interactions can bemodelled mathematically). When a morphogen blocks anther signal, we call that antagonism, and antagonist signals are just as common as induction signals. They must, after all, it is fundamentally just as important to grow a head as it is to not grow two heads.
Proliferation
Growth of a tissue is also called proliferation. Proliferation occurs either by mitosis (adding cells) or by adding extra-cellular matrix (synthesizing proteins or other molecules). When new growth happens next to a region of older growth, this is appositional growth, whereas if growth happens from within and pushes older tissue outwards, that is interstitial growth. Dense tissues, such as bone, enamel and dentin, undergo appositional growth. Softer tissues like mesenchyme grow interstitially.
The growth of a tissue is regulated by hormones, such as Growth Hormone, which coordinates the rate of growth of tissues body-wide. On the other hand, local-actingmorphogens cause small regions to grow faster than neighboring regions. To make room for new cells while still maintaining a single-layer of cells connected to their neighbors by desmosomes and tight junctions, an epithelium bulges inwards or bulges outwards if it can't grow wider. An outward growth is known as a bud, and an inwards growth is an invagination. Arms and legs grow from buds, while teeth, hairs and the brain grow from invaginations. These are both forms of interstitial growth.
Differentiation
Differentiation is the process where stem cells begin to express different genes, causing them to appear and function differently from other cells. All cells in the human body have the same DNA, because all cells copied their DNA from the DNA in the zygote(although a few exceptional cells do change their DNA a bit). The zygote is a toti-potent stem cell, transcription factors could potentially bind to and activate any gene. As cells differentiate, however, they begin to limit their cell fate by packing certain genes by methylating them and packing them around histones.
Part of the differentiation process that follows receiving a morphogen signal, target cells methylate un-needed genes, permanently shutting those genes off. This limits the fate of these cells. After that occurs the morphogen is no longer needed, because the methylation pattern is copied during mitosis and passes to both daughter cells.
In the image to the left, red morphogen instructs one cell to methylate green genes, leaving the red genes open to potentially be transcribed. The green morphogen does the opposite to a different cell. As the red cell duplicates its DNA, it copies the methylation pattern, so that all of its daughter cells will not be able to express green genes. Meanwhile, the green cell produces a number of daughter cells who share the green methylation pattern, none of its daughter cells will express red genes. We now have 4 red cells, who belong to one lineage, and 4 green cells who share a different lineage. Further patterning arises as cells transcribe un-methylated genes. In the cartoon, red cells transcribe a new short-range morphogen gene, which induces nearby green cells to transcribe the dark-green gene. We would say those target cells have differentiated further, and this organism has become more complicated. Generally speaking, as a cell bomes more specialized, it's fate becomes more limited, possibly until only one fate is possible.
Morphogenesis is the appearance of new shapes in an organism. It means from no shape to some shape. Morphology is the study of these shapes. From the zygote to the blastula stage (sometimes referred to as the germinal stages) the ball of cells is amorphous, it has no shape (other than being a sphere and not a cube or sausage). At the start of gastrulation, however, the embryo morphs (transforms its shape). New shapes arise in the developing embryo. It develops an inside and an outside, a tail end and a head end, soon it will develop a left and right, and a front and back. After that, a nervous system and a circulatory system become visible. How long do you think it will take before teeth develop? The answer may surprise you.
The image I have used here is a famous drawing of tiny sea creatures with mineralized exo-skeletons. This image was later analyzed by a mathematical biologist named D'Arcy Wentworth Thompson, who noticed different biological shapes can be different transformations of the same shape (transformations, he proposed, occured by different localized rates of growth). He is regarded as the first person to describe morphogenesis. When Thompson looked at the tiny sea creatures, he didn't see multiple shapes, he saw changes to one basic shape. These changes happened over evolutionary time, and led to different sea creatures (who shared a single lineage). In this book, we looks at different body parts that share the same lineage but change over developmental time, such as hair follicles and tooth buds. Thompson's ideas were expanded upon by the mathematician Alan Turing, whose other achievements include inventing computer science and breaking the Nazi code during WWII. Sadly, he committed suicide after his arrest, conviction and imprisonment for homosexuality by the British government he helped save. Their government's view on homosexuality has developed since then, we can understand the current view better by following its lineage.Generalized scheme of embryonic development, by JMW Slack is liscensed CC BY SA 4.0
Maturation
Maturation is the process by which new shapes created by morphogenesis continue to develop and take on a more mature form. By the end of week 9, all of the basic organ shapes will have formed, and we will say the embryonic period is over and the fetal stages begin. The fetus will undergo maturation, and maturation will involve bits of all the previous processes.
In the construction of tall buildings or large ships, scaffolding is built first to allow the contruction workers to get where they need to, then removed when construction is complete. Similarly, during development, apoptosis (programmed cell death) is an integral part of the construction of body parts. Many tissues grow more cells than are needed in the mature form. To undergo maturation, un-needed cells undergo apoptosis. For instance, during the growth of bone tissue, extra cartilage tissue is made to act as a scaffold for osteoblasts. Shown in the image here, the hands and feet start of as amorphous paddles, and only with apoptosis in alternating regions will the morphogenesis of fingers and toes occur. Without enough apoptosis, people can be born with webbed fingers or toes, or suffer ankyloglossia (their tongue is rigidly anchored to the floor of the mouth by a pronounced lingual frenulum).
The pre-implantation period begins with fertilization, spans a week or so of development, and ends with an embryo implanting into the uterus. Fertilization is the union of one sperm cell and one egg-- thousands of sperm are needed, but only one is allowed into the egg. This union joins 23 maternal chromosomes with 23 matching paternal chromosomes, forming a zygote. Fertilization most often occurs towards the end of a fallopian tube. Over the next week the zygote will undergo mitosis and increase in cell number, but it will not grow in size. All of the amino acids and nucleotides for making new cells during this first week come from the cytoplasm of the zygote, which is why the egg is a very large cell. After implantation, the uterus will provide the raw materials for an embryo to grow larger.
Non-disjunction of sister chromatids, by Wpeissner is liscensed CC BY 3.0
During mitosis, the 23 pairs of chromosomes are duplicated and divided between 2 daughter cells evenly. If mistakes are made and an odd number of chromosomes wind up in a daughter cell, this fails a cell cycle checkpoint and triggers apoptosis. If a chromosome breaks, it can lead to the same result.
Boy with Down Syndrome using cordless drill to assemble a book case, by Rob Kay is liscensed CC BY SA 3.0
There are exceptions, however. A cell with an odd nuber of chromosome 21 or 23 can survive. If that cell is the egg, then the zygote and all subsequent cells will have an odd number of chromosomes. This leads to congenital disorders such as Trisomie 21. If the chromosomal segregation error happens at the 2-cell stage of development, than only half of the cells in the adult would have an extra chromosome, and the condition would be less severe. Extra chromosomes interfere with decision-making processes in development. People with Trisomie 21 frequently develop microdontia, and may suffer from hyposalivation and bruxism. They therefore need extra care maintaining healthy oral hygiene.
Early divisions
During the first week of development, the zygote will undergo 3 cleavage divisions, where mitosis occurs in a synchronized fashion, and all the cells divide along a single plane, producing an 8-cell embryo. Synchronizing mitosis requires cell-to-cell communication between gap junctions to coordinate cell cycle checkpoints. Synchronized mitosis leads to the embryo growing evenly in all directions. The fact that multiple cells are dividing in the same direction means these cells have some direction (or polarity), even if the embryo as a whole has no visibe top or bottom yet.
As mitosis continues, the number of cells continues to double, but the cells are not dividing along the same plane. This solid ball of cells is called a morula. As mitosis continues, the cell count doubles and the cells continue to get smaller, until the solid ball of cells becomes a hollow ball of cells named a blastula. From the zygote to the blastula stage, none of the cells appear any different, their only changes are in size. A fancier way to say this is that there are no changes to morphology.
The blastula stage offers what could be (but isn't) considered the first morphological change. A group of cells named the embryonic disc move away from the others. The fate of the embryonic disc is to become the embryo, while the outer cells, known as the trophoblast, are fated to become extra-embryonic structures such as the placenta. The blastula will implant in the endometrium about a week after fertilization and continue developing.
Gastrulation
Gastrulation: cross-sectional view
Gastrulation overview
It is not birth, marriage, or death, but gastrulation, which is truly the most important time in your life.--Lewis Wolpert.
We left off with a hollow ball of identical cells, the blastula. During gastrulation, some of these cells invaginate, forming a layer of cells on the inside and a layer of cells on the outside. The significance of this is that this is the first instance where embryonic cells become different from one another. But, wait, if all the blastula cells are indentical, why did only some of the gastrulate? Mitosis does indeed generate 2 cells with identical DNA, but not necessarily identical cytoplasm. If an egg had mRNA for a transcription factor clustered into one spot in the cytoplasm, the first cleavage division would produce two non-identical cells, one with the transcription factor mRNA and one without. Later, when the mRNA was translated, it could induce that cell to migrate inwards, while other cells stayed on the outside. This is known as a maternal effect, because the mRNA for the trancription factor was not transcribed from the egg's DNA, but was made by mom's ovary cells and deposited into the egg. The end result of gastrulation is an embryo with two layers (outside and inside). The tube that forms will become the gastrointestinal tract, and that opening-- can you guess-- will become the anus. The embyro therefore now has a distinct tail direction and head direction. There is an old developmental biology joke, and it's funny because it's true: at one time in your life, you were nothing but an asshole.
The image I have drawn represents the way a sea urchin gastrulates. Human embryos look different because it is only the embryonic disc that undergoes gastrulation, not the entire blastula. But the 2 layers are the same, human gastrulas simply look more squashed. Oh, and sea urchins don't form anus-first. The reason sea urchin gastrulation looks simpler than human gastrulation is that human blastulas reserve some cells (the trophoblast) to form a placenta and amnion, while sea urchins dump their eggs into the sea.
Gastrulation: surface viewNext, cells from the outer layer migrate between the two layers. This gives the embryo 3 layers, which are the 3 basic germ layers. The outer layer of ectoderm and the inner layer of endoderm remain epithelial in appearance, while the middle layer of cells (mesoderm) underwent a morphological change, and resembles mesenchyme. What's more, this migration begins at a single location called Hensen's node, which extends down the embryo forming the primitive streak. The side of the embryo with the node is fated to become the dorsal side, and the streak divides the embryo into a left-half and a right-half. You have now witnessed, albiet in cartoon form, how something (all the basic body directions) arose from nothing. Don't let the significance of this sneak past un-noticed. Scientists have a hard time explaining what there was before the universe formed, or how the Earth went from having no life to having life. Embryologists, by contrast, explain how embryos change from having no shape to having shape in great detail (we've had the benefit of directly watching and manipulating this process).
3 embryonic layers
Cell fate
Ectoderm
Epithelium of skin and oral mucosa, neural tissue
Mesoderm
Connective & muscle tissue
Endoderm
Epithelial lining of hollow organs
Epithelial-to-mesenchymal transition
The process by which ectodermal cells quit being ectoderm and migrate to the middle and form mesoderm is called an epithelial-to-mesenchymal (EMT) transition. The epithelial cells of the ectoderm must first lose their cell-to-cell contacts with neighboring cells, which makes them less-epithelial. They lose their polarity and become mesenchymal stem cells. This process is not only required to form mesoderm during gastrulation, it also occurs during neural crest migration, wound healing, and cancer metastasis. Cells in the mesoderm (the name for the middle layer) form mesenchyme (a type of tissue), while the other two layers remain epithelial.
There is a reverse process, called a mesenchymal-to-epithelial (MET) transition, which also occurs during wound healing. Together, EMT allows epithelial stem cells from a healthy region of oral mucosa to turn into mesenchymal stem cells, migrate away from the healthy region into a damaged region. MET then allows the mesenchymal stem cells to turn back into epithelial stem cells, which divide and differentiate into new keratinocytes. Because this mimics what happens during gastrulation, we say "wound healing recapitulates (states again) development".
Differentiation of ectoderm
Neurulation
Gastrulation gave the embryo its polarity, as well as the beginning of the GI tract. One of the next organ systems to develop is the central nervous system. The basic processes used in neurulation are re-used when other hollow organs form. First, underlying mesoderm known as the notochord sends morphogens to nearby ectodermal cells. This induces the nearby region of ectoderm to differentiate into neuro-ectoderm, followed by local proliferation. Because these epithelial cells cannot spread side-to-side they invaginate. Ultimately, invaginating cells lose their contacts with the ectoderm and fuse together, creating a seperate structure called the neural tube. This tube will develop into the brain and spine.
The neural tube begins growth from the anterior portion of the embryo, then zips up in an anterior-to-posterior direction. The zippering is not perfect, as the anterior end has to grow a bit extra before it zips up to form the primitive brain. For all of the developmental processes we will and won't be covering, the anterior end is usually the region that develops first, followed by more posterior regions. Zipping the neural tube requires adequate levels of folic acid, so it is important that mother's melanin has protected this vitamin, otherwise incomplete closure of the neural tube may result.
Some neuro-ectoderm cells do not fuse together. Instead, these cells undergo an epithelial-to-mesenchymal transition and migrate away from the neural tube. These cells are called neural crest cells, and they are fated to become a number of important cells throughout the body, including melanocytes, odontoblasts, dental pulp, cementoblasts, and the ecto-mesenchyme of the pharyngeal arches. Even after they migrate to new and distant tissues, they retain signs of their lineage, such as the dendrites of melanocytes. To make migration easier, neural crest cells express an enzyme called a matrix metalloprotease (MMP). This enzyme digests proteins found in the extra-cellular matrix. By now I hope you are tired of me saying wound healing recapitulates development, but this enzyme is re-used to heal certain type of tooth wounds.
Plate 20, by henrey Grey is in the Public Domain CC0
Somite formation
During the same time as neurulation, the mesoderm undergoes differentiation. Mesoderm starts off as an amorphous layer of mesenchyme, but regions of mesenchyme pinch off in repeating spherical structures called somites, forming segments along the anterior-to-posterior axis of the embryo. The fate of the somites is to become solid organs, either repeating units of connective tissue such as the ribs and vertebrae, or repeating units of muscle tissue such as the rectus abdominus and intercostal muscles. The division of each somite involves a mesenchymal-to-epithelial transition, as some mesenchymal stem cells differentiate into an epithelium that separates one somite from the rest of the mesenchyme.
In the third week, mesoderm also begins to develop into blood, the heart and the circulatory system. Mesenchymal stem cells first differentiate into blood islands. Next, angiogenesis (the growth of new blood vessels) begins. Two larger blood vessels will fuse-- with some help from neural crest cells to form the primitive heart, which begins beating.
Differentiation of endoderm
Formation of the pharynx
The endoderm that migrated interiorly during gastulation extends its way towards the anterior end of the embryo. This forms the primitive foregut, which lter becomes the pharnynx, oral and nasal cavities. At this time, however, there are no openings for the mouth and nostrils.
You, negative 8 months old
4th week
We are now only up to the 4th week of development. A lot has happened! At this time, the embryo continues to undergo local regions of proliferation, pattering, fusion and the other processes covered at the beginning of the chapter. This leads to the formation of rudimentary (basic) structures. When a structure first becomes visible from the surface of an embryo, it is often called a placode (an ectodermal thickening). The cells are fated to become an organ, but at this time it is shape is no more than a raised bump or shallow pit. By the 4th week, the following organs begin to develop:
Structure
fate
Eye placodes
primitive eyes
Otic placodes
primitive ears
Nasal placodes
primitive nose
Pharyngeal arches
so many things that these get their own chapter
Oro-pharyngeal membrane
primitive mouth (not open yet)
Clinical applications of early development
Disturbances in these early stages of development are usually so severe the embryo does not survive. However, a few clinical conditions exist related to these early stages and to oral health.
Ectodermal dysplasia, by Pratheeba227 - photographed at GOVT VELLORE MEDICAL COLLEGE, is liscensed CC BY SA 3.0
Ectodermal Dysplasia
Problems with the induction of neural crest cells during neurulation leads to disturbances in the formation of teeth, hair follicles, salivary glands and other structures. What these have in common is they are all specialized structures of the ectoderm, induced by neural crest cells to differentiate. Before they form, neuro-ectoderm cells must receive the correct morphogens, which in turn must activate or inactivate the correct transcription factors to trigger differentiation into neural crest cells. Some neural crest cells migrate to distant regions of ectoderm, determine their location by interacting with morphogens in the ground substance, and release different morphogens to induce regions of ectoderm to differentiate into sweat glands, salivary glands, teeth or hair follicles. Mutations in any of the genes for these morphogens, their receptor proteins, or down-stream transcription factors may lead to a condition named Ectodermal Dysplasia. This group of syndromes is rare, with only 7000 cases worldwide, but there are over 150 different gene that, when mutated, can cause this condition. For instance, mutations that interfere with a Wnt morphogen, which is required for neural crest cell induction, reduces the number of neural crest cells, which are in turn needed to induce the formation of hair follicles, tooth buds, sweat glands, and many other structures in the human body. People with Ectodermal Dysplasia often have fewer teeth and smaller, under-formed teeth (hypodontia), and may also suffer from xerostomia. Dental implants or dentures can be part of the treament for this condition, as well as the use of artificial saliva and tears.
Cleido-cranial dysostosis (CCD) is a congenital disorder caused by a mutation to a transcription factor required for the differentiation of bone and teeth. It is required to trigger osteo-chondro-progenitor cells to exit the cell cycle and differentiate into osteoblasts. It is also re-used to induce the differentiation of odontoblasts. After teeth have formed, this transcription factor is re-used once again to activate the expression of a Matrix metalloprotease enzyme, which is necessary for remodeling of the alveolar sockets. Without this enzyme, retention of deciduous teeth occurs. Dental implants or dentures (such as the ones the actor and CCD-philanthropist Gaten John Matarazzo III have in this photo) are the preferred treatment. In addition, a person with cleido-cranial dydostosis may have small clavicles and changes to shape of the skull-- bones that form by intra-membranous ossification.
Figure 3, by Beatriz A. Ro isdas-Junco at al, is liscensed CC BY 3.0
Stem cell therapies
When cells differentiate, they permanently inactivate un-needed genes by methylation and storage around histones. Researchers are learning ways to reverse this process, and cause differentiated cells to revert to a stem-cell state. This raises the possibility of promoting regeneration of tissues that do not otherwise regenerate, removing the need for tissue grafting or transplantation. Because many dental tissues do not regenerate well, there is potential for these technologies to be applied to the oral cavity, such as growing biological dental implants instead of using metals and ceramics.
However, more interest has been placed on acquiring mesenchymal stem cellsfromdental tissues. For instance, stem cells isolated from maxillary third molars have been used in clinical trials to improve healing and reduce the need for transplanted tissue in maxilofacial surgery. But because of their potential to differentiate into a wide array of cells, great interest has been placed on collecting dental stem cells to treat diseases unrelated to the oral cavity. With the correct morphogen and plenty of dental stem cells, it may be possible to reverse the damage caused by neurodegenerative disease such as Alzheimer's Disease (AD) and Parkinson's Disease (PD), spinal trauma, myocardial infaction (heart attacks), and Muscular Dystrophy (MD). What else would you do with extracted third molars?
Unfortunately, there is no legal definition here in the U.S. for what constitutes safe, effective stem cell therapy. Even if good rules are put in place, medical-tourism will not abide. In addition to honest and good research in stem cell therapies (example), there are also a large number of dubious-to-downright-quack stem cell clinics operating in the grey market, using risky, untested methods and making absurd claims to take as much money as possible from vulnerable people. So who's who? I think, in general, therapies that use or boost a patient's own stem cells have the best possible chance of providing positive outcomes with minimal risk of side-effects-- especially if a therapy appears seems to be targetted, such as by adding a specific morphogen to boost a specific result. On the other hand, simply throwing stem cells at a problem-- cells from other people, other animals, or from non-animal sources-- carries the biggest risk of serious adverse effects with minimal benefits. Everyone loves cutting-edge technology, myself included. Keep in mind that 19 out of 20 (or so) potential drugs that are demonstrated scientifically to work in animal models fail human trials. That means there is plenty of scientific evidence out there for treatments that were later shown to not work, but people can still cherry-pick the positive information and use it to sell a product. I think its best to mix our optimism for fascinating new technologies with good doses of caution and patience.